Схема строения возбудителя герпеса

Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 19 февраля 2019;
проверки требуют 5 правок.
Герпесви́русы[3] (лат. Herpesviridae) — большое семейство ДНК-содержащих вирусов, вызывающее разнообразные болезни не только у человека и других млекопитающих, но и у птиц, рептилий, амфибий, рыб. Герпесвирусами болеют большинство населения нашей планеты[4].
Общие сведения[править | править код]
По состоянию на май 2016 года в Международном комитете по таксономии вирусов (ICTV) зарегистрированы 86 видов[2]. Отличительным признаком вирусов этого семейства является нахождение вируса в клетках латентно, персистируя, бесконечно длительное время, без клинических проявлений.
Персистенция (вирусов) (лат. persistere — оставаться, упорствовать) — термин, предложенный в 1923 году французским бактериологом, иммунологом и вирусологом Константином Левадити (Levaditi Constantin, 1874—1953) и румынским вирусологом Штефаном Николау (Nicolai Stefan, 1896—1967), обозначающий длительное сохранение вируса в организме хозяина или в клеточной культуре.
Название семейства происходит от греч. ἕρπειν (herpein) ‘ползать’ через латинское herро ‘ползаю’. Инфекционные болезни, вызванные вирусами этого семейства, протекают остро, переходят в латентную стадию, не проявляют себя до определённого времени.
При ухудшении состояния организма (простуда, стресс, утомление и т. п.) вирус даёт о себе знать болезнью. Примером может служить герпетические высыпания на губах при «простуде».
Вирусы семейства Herpesviridae имеют общие биологические свойства. Они имеют эффективные механизмы взаимодействия с иммунной системой хозяина, позволяющие им достичь максимального распространения и сохраняться в организме в течение всей жизни.
В ходе своей жизнедеятельности вирусная ДНК экспрессирует определённые группы генов и, соответственно, кодируемых ими белков, которые, собственно, и определяют жизненный цикл вируса в клетках хозяина, приводя к изменению их фенотипических свойств, то есть трансформации.
Трансформация клеток вызывает развитие определённых иммунопатологических реакций, направленных против собственного организма и приводящих к вирусиндуцированной иммуносупрессии и длительной персистенции вируса в организме человека. В клетках своего хозяина они переходят в латентное состояние. При латентном состоянии нарушается полный репродуктивный цикл вируса. Он находится в клетках хозяина в виде субвирусных структур.
Клинические формы герпесвирусных инфекций характеризуются выраженным полиморфизмом. Существуют различные формы проявления герпесвирусных инфекции.
В любом случае герпесвирусы остаются в инфицированном организме пожизненно. Вирус может периодически рецидивировать с характерными клиническими проявлениями или бессимптомно, или приобретать генерализованный характер с возможным летальным исходом.
Строение[править | править код]
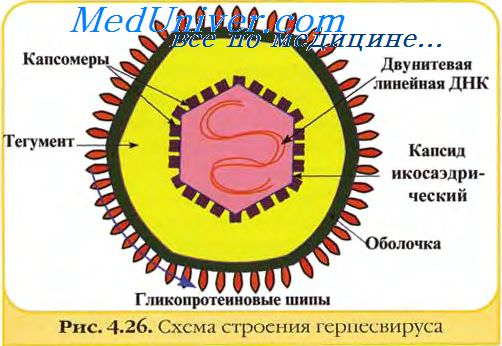
Вирионы семейства вирусов герпеса имеют сферическую форму диаметром от 120 до 300 нм. Внутри зрелой вирусной частицы содержится 35—45 различных белковых молекул.
В центре вириона различают сердцевину (кор, от лат. cor) размером 75 нм, содержащую ДНК.
Эта сердцевина окружена оболочкой — капсидом. Размер капсида составляет 100—110 нм. Он построен из множества одинаковых геометрически правильных белковых структур — капсомер. Капсомеры точно подобраны и подогнаны друг к другу, образуя двадцатиугольник, по-гречески икосаэдр. Капсид содержит 162 капсомера: 150 шестиугольных (гексамеров) и 12 пятиугольных (пентамеров).
Вокруг капсида имеется аморфный белковый тегумент и всё это заключено в оболочку с гликопротеиновыми шипами.[источник не указан 2084 дня]
Геном[править | править код]
Геном герпесвирусов представляет собой линейную двухцепочечную молекулу ДНК. У герпесвируса человека 1-го типа она имеет размер 152 261 пара нуклеотидов и содержит 77 генов, все из которых кодируют белки; доля пар ГЦ составляет 68 %[5]. Геном герпесвируса человека 2-го типа состоит из 154 746 пар нуклеотидов и несёт 77 генов; доля пар ГЦ составляет 70 %[6].
Механизм инфицирования[править | править код]
Герпесвирусами заражаются человек и животные.
Образование новых вирусных частиц идёт под управлением вирусного генома. При попадании в организм носителя герпесвирусы адсорбируются клетками-мишенями на клеточной поверхности и освобождаются от капсида и дополнительной оболочки-конверта.
После эндоцитоза вирусной сердцевины происходит интегрирование вирусной ДНК с ядерным материалом клетки. Затем на ядерной мембране происходит образование и созревание новых вирионов и последующее их отпочковывание клеткой посредством экзоцитоза.
Для оболочки вируса, его капсида и ДНК, используются аминокислоты, белки, липопротеиды, нуклеозиды клетки-хозяина. По мере истощения внутриклеточных резервов, эти молекулы поступают в инфицированную клетку из межтканевых пространств.
Для культур, хронически зараженных вирусом простого герпеса, характерно[7]:
- возникновение состояния носительства после деструкции основной массы клеток и репопуляции немногих выживших;
- постоянное наличие очагов дегенерации, морфологически характерных для этого вируса;
- небольшой процент инфицированных клеток;
- низкая продукция вируса одной инфицированной клеткой;
- отсутствие интерферона в среде
- определенная резистентность клеток, освободившихся от состояния носительства, к реинфекции гомологичным вирусом.
В природе насчитывается восемь типов вирусов из семейства Herpesviridae, вызывающих болезнь у человека.
Классификация[править | править код]
Семейство герпесвирусов по классификации Международного комитета по таксономии вирусов (ICTV) делится на подсемейства. Используются критерии:
- структура генома вируса
- белковый состав
- характер репликации вируса
- среда носительства
- тропизма к тканям
- распространение вируса в культуре
- длительности репродуктивного цикла
Под этим признаком различают подсемейства:
- Alphaherpesvirinae — альфагерпесвирусы, или α-герпесвирусы
- Betaherpesvirinae — бетагерпесвирусы, или β-герпесвирусы
- Gammaherpesvirinae — гаммагерпесвирусы, или γ-герпесвирусы
- 1 вид, не входящий ни в одни подсемейство и род (incertae sedis)
В 2016 году всем видам семейства изменили научное название для улучшения номенклатуры видов, при этом существенно не меняя названия герпесвирусов человека — к слову virus добавили префикс (alpha-, beta-, gamma-), указывающий подсемейство, к которому этот вид относится[8].
Герпесвирусы человека[править | править код]
Выделяют восемь типов вирусов герпеса, вызывающих разные по тяжести процесса заболевания у людей. Характерной особенностью заболеваний является нахождение вирусов в организме человека в латентном состоянии[9][10].
| Вид герпесвируса человека[3][2] | Название вируса | Подсемейство вирусов | Род вирусов | Вызываемая болезнь |
|---|---|---|---|---|
| Герпесвирус человека тип 1 (ГВЧ-1, Human alphaherpesvirus 1) | Вирус простого герпеса первого типа (ВПГ-1, Herpes simplex virus-1, HSV-1) | Альфагерпесвирусы | Simplexvirus | Оральный и генитальный герпес, но чаще оральный (герпетический стоматит, губной герпес) |
| Герпесвирус человека тип 2 (ГВЧ-2, Human alphaherpesvirus 2) | Вирус простого герпеса второго типа (ВПГ-2, Herpes simplex virus-2, HSV-2) | Альфагерпесвирусы | Simplexvirus | Оральный и генитальный герпес, но чаще генитальный и вагинальный герпес |
| Герпесвирус человека тип 3 (ГВЧ-3, Human alphaherpesvirus 3) | Вирус ветряной оспы (Varicella-zoster virus, VZV) | Альфагерпесвирусы | Varicellovirus | Ветряная оспа (варицелла, varicella), опоясывающий лишай (зостер, zoster) |
| Герпесвирус человека тип 4 (ГВЧ-4, Human gammaherpesvirus 4) | Вирус Эпштейна — Барр (ВЭБ, Epstein-Barr virus, EBV) | Гаммагерпесвирусы | Lymphocryptovirus | Инфекционный мононуклеоз, лимфома Беркитта, лимфома ЦНС у больных с иммунодефицитным синдромом, посттрансплантантный лимфопролиферативный синдром (post-transplant lymphoproliferative syndrome, PTLD), назофарингеальная карцинома |
| Герпесвирус человека тип 5 (ГВЧ-5, Human betaherpesvirus 5) | Цитомегаловирус человека (ЦМВ, Human cytomegalovirus, HCMV) | Бетагерпесвирусы | Cytomegalovirus | Инфекционный мононуклеоз, ретинит, гепатит, увеличение органов брюшной полости, воспаление слюнных желез (так называемое слюнотечение) |
Герпесвирус человека тип 6 (ВГЧ-6):
| Розеоловирусы:
| Бетагерпесвирусы | Roseolovirus | Шестая болезнь — детская розеола (розеола инфантум, roseola infantum) или экзантема (экзантема субитум, exanthem subitum) |
| Герпесвирус человека тип 7 (ВГЧ-7, Human betaherpesvirus 7) | Розеоловирус (Human herpesvirus 7, HHV-7) | Бетагерпесвирусы | Roseolovirus | Вероятная причина синдрома хронической усталости (СХУ)[3]. Часто сосуществует с вирусом герпеса 6 типа |
| Герпесвирус человека тип 8 (ВГЧ-8, Human gammaherpesvirus 8) | Герпесвирус, ассоциированный с саркомой Капоши[en] (Kaposi’s sarcoma-associated herpesvirus, KSHV) | Гаммагерпесвирусы | Rhadinovirus | Саркома Капоши, первичная лимфома серозных полостей, некоторые разновидности болезни Кастельмана |
Примечания[править | править код]
- ↑ Таксономия вирусов (англ.) на сайте Международного комитета по таксономии вирусов (ICTV).
- ↑ 1 2 3 Таксономия вирусов (англ.) на сайте Международного комитета по таксономии вирусов (ICTV). (Проверено 30 июня 2016).
- ↑ 1 2 3 Атлас по медицинской микробиологии, вирусологии и иммунологии : Учебное пособие для студентов медицинских вузов / Под ред. А. А. Воробьева, А. С. Быкова. — М. : Медицинское информационное агентство, 2003. — С. 109. — ISBN 5-89481-136-8.
- ↑ Вирусы Герпеса. Герпесвирусная инфекция
- ↑ Human herpesvirus 1, complete genome
- ↑ Human herpesvirus 2, complete genome
- ↑ Анджапаридзе О. Г., Богомолова Н. Н. Моделирование и исследование хронических форм вирусных инфекций в культурах клеток. — Москва: Медицина, 1974. — С. 161. — ISBN УДК 616.988-036.12-092.4.
- ↑ Davison A., Pellett P., Stewart J. Rename species in the family Herpesviridae to incorporate a subfamily designation : [англ.] // ICTVonline. — Code assigned: 2015.010aD. — 5 p.
- ↑ Whitley R. J. Herpesviruses. in: Baron’s Medical Microbiology (Baron S et al., eds.) (англ.). — 4th. — Univ of Texas Medical Branch, 1996. — ISBN 0-9631172-1-1.
- ↑ Murray P. R.; Rosenthal K. S.; Pfaller M. A. Medical Microbiology (неопр.). — 5th. — Elsevier Mosby, 2005. — ISBN 978-0-323-03303-9.
Источник
Эпидемиология герпесвирусов. Строение герпесвирусов.Герпесвирусы могут передаваться от одного поколения хозяев к другому благодаря персистенции, часто латентной инфекции, при которой вирус периодически реактивируется и выделяется из организма. При некоторых герпесвирусных инфекциях выделение вируса может быть фактически непрерывным. Молекулярное филогенетическое изучение подтверждает, что, за небольшими исключениями, каждый вирус является уникальным, потому что он происходит от своего видового хозяина. Латентность позволяет герпесвирусам быть пожизненными даже в очень небольшой изолированной группе хозяев. Попытки разработать критерии классификации герпесвирусов внутри семейства пока не увенчались успехом. Существующая классификация основана на их естественном паразитизме. В зависимости от биологических свойств герпесвирусы делятся на 4 подсемейства, 3 из которых имеют названия альфа-, бета-, гаммагерпесвирусы и одно — без названия, объединяющие герпесподобные вирусы. Подсемейство альфагерпесвирусы включает герпесвирусы 1 и 3 человека, герпесвирус 1 кур и вирус болезни Марека. Большинство альфагерпесвирусов размножаются быстро (цикл репликации меньше 24 ч) и вызывают латентную инфекцию первоначально в сенсорных ганглиях. Подсемейство бетагерпесвирусы представлено цитомегаловирусами многих видов млекопитающих. Прототипным вирусом является герпесвирус человека 5 — цитомегаловирус человека. Вирусы этого подсемейства размножаются медленно (цикл репликации более 28 ч) и вызывают лизис клеток спустя несколько дней. Они могут оставаться латентными в секреторных железах, лимфоретикулярных тканях, почках и других органах. Подсемейство гаммагерпесвирусы представляет лимфотропные вирусы. Прототипом является герпесвирус человека 4 — вирус Эпштейн-Барр. Гаммагерпесвирусы обладают узким хозяинным спектром и находятся латентно в лимфоцитах (лимфома Беркитта и др.), некоторые представители вызывают инфекцию с разрушением клеток эпителия и фибробластоидных клеток. Вирионы герпесвирусов имеют диаметр около 120—150 нм и состоят из 4 структурных компонентов: сердцевины (ядра), содержащей линейную двуцепочечную ДНК размером 125—235 тпн, икосаэдрического нуклеокапсида диаметром 100-110 нм, содержащего 162 капсомера; аморфного материала (тегумент), окружающего капсид и оболочки с небольшими гликопротеиновыми выступами (пепломерами) на поверхности.
ДНК-геном закручен вокруг волокнистого ядра, имеющего вид катушки, которая прикреплена волокнами к внутренней поверхности окружающего капсида. Вирусный геном в соответствующих экспериментальных условиях обладает инфекционностью. Сердцевина вириона окружена икосаэдрическим капсидом, содержащим 162 частично полых капсомера (12 пентамеров и 150 гексамеров). Капсомеры высотой 12,5 нм и диаметром 9,5 нм имеют внутренний осевой канал диаметром 4 нм, простирающийся внутрь на половину длинной оси капсомера. Слой, лежащий между оболочкой вириона и нуклеокапсидом (тегумент), содержит глобулярный материал, часто распределяемый ассиметрично, и выражен неодинаково у разных герпесвирусов. У вируса герпеса простого он развит слабо, тогда как у вирусов болезни Марека (БМ) и чумы уток (ЧУ) он развит отчетливо и его наличие обусловливает крупный размер вирионов. На поверхности герпесвирусов находится двухслойная оболочка с многочисленными небольшими гликопротеиновыми пепломерами, образованными главным образом за счет «бляшек» измененной ядерной мембраны инфицированной клетки. Количество гликопротеинов в оболочке герпесвирусов варьирует. В оболочке вируса простого герпеса их не менее 11. Число копий индивидуальных гликопротеинов может превышать 1000 на вирион. Содержание полипептидов в вири-оне точно не известно и различается у разных герпесвирусов. В общем, оно колеблется в пределах 35—45 полипептидов.
Все герпесвирусы кодируют широкий набор ферментов, связанных с метаболизмом нуклеиновых кислот, синтезом ДНК и процессингом белков. Синтез вирусных ДНК и сборка вирионов происходят в ядре клетки. Капсиды покрываются оболочкой, проходя через мембрану ядра. Образование инфекционного вирусного потомства сопровождается деструкцией зараженных клеток. Вероятно, что некоторые кодируемые вирусом белки играют заметную роль в патогенезе и иммунитете при герпесвирусной инфекции, не являясь необходимыми для репликации вируса в культуре клеток. Считают, что геномы герпесвирусов в латентном состоянии в клетках хозяина существуют в циркулярной эписомальной (экстрахромосомальной) форме. Репликация герпесвирусов наиболее детально изучена на примере герпесвируса 1 человека (вирус простого герпеса 1). После прикрепления вириона путем связывания пепломерных гликопротеинов оболочки с рецепторами клетки, одним из которых является гепаринсульфатпротеогликан, нуклеокапсид входит в цитоплазму путем слияния оболочки вириона с мембраной клетки путем эндофагоцитоза. Затем ДНК-белковый комплекс освобождается от нуклеокапсида и быстро останавливает синтез макромолекул клетки. Геном ВПГ-1 (153 тпн) содержит три области инициации-репликации. Он практически кодирует все гены, необходимые для репликации, кроме ДНК-лигазы и топоизомеразы. На первом этапе репликации функционирует ДНК-полимераза а, а затем функционирует вирусная ДНК-полимераза. Три класса мРНК — а, (3 и 7-транскрибируются последовательно клеточной РНК полимеразой II. Таким образом а (ранние) РНК в последующем функционируют как мРНК и транслируются в а-белки, которые вызывают транскрипцию (3 (ранних) мРНК, трансляция которых приводит к синтезу (3 (ранних) белков и подавляет дальнейшую транскрипцию а мРНК. Затем начинается репликация вирусной ДН К с использованием тех же самых аир протеинов, так же как протеинов клетки. Транскрипционная программа затем возобновляется, и в результате появляются 7 (поздние) мРНК, которые транслируются в 7 протеины. Более 70 белков, кодируемых вирусом, образуются в цикле его репродукции, многие из а и (3 белков являются ферментами и белками, связанными с вирусной ДНК, тогда как 7 белки являются структурными вирионными компонентами. Сложный контроль регуляции экспрессии осуществляется на уровне транскрипции и трансляции. Вирусная ДНК реплицируется в ядре, и вновь синтезированная ДНК упаковывается в преформированные незрелые капсиды. Созревание вирионов связано с вхождением ДНК в нуклеокапсиды и связыванием нуклеокапсидов с измененным внутренним слоем оболочки ядра и последующим обволакиванием вирионов при почковании. Зрелые вирионы накапливаются внутри вакуолей в цитоплазме и освобождаются экзоцитозом или при цитолизе. Вирусспецифические белки обнаружены также в цитоплазматической мембране, где они участвуют в клеточном слиянии и могут в качестве Fc-рецепторов выступать мишенями в иммунном цитолизе. Герпесвирусная инфекция сопровождается образованием внутриядерных телец включений, которые обычно обнаруживают в фиксированных окрашенных препаратах. Геном вируса кодирует синтез по меньшей мере четырех гликопротеинов (gB, gC, gD, gE). Имеет место посттрансляционное расщепление или другие модификации вирусных белков. Белок gB обладает фузогенной активностью и ответственен за проникновение вируса в клетку; gD — основной протективный антиген. Белки gC и gE не играют существенной роли в репродукции вируса, но помогают ему ускользнуть от иммунной системы организма. В отсутствие репликации вирус редко вызывает сплавление клеток, а при наличии специфических условий — агглютинацию эритроцитов. Оболочка вириона играет существенную роль в антигенности и инфекционности вируса. Гликопротеины вирусной оболочки ответственны за прикрепление вирусной частицы к клетке, проникновение вируса в клетку, за индукцию нейтрализующих антител и образование иммунитета. Нейтрализующие антитела реагируют с главными вирусными гликопротеинами, находящимися в оболочке вириона. Мембранные антигены, индуцированные в зараженных клетках, содержат по крайней мере три гликопротеина, обнаруженных в структуре вириона. Эти данные послужили основанием для приготовления вакцин из вирусспецифических компонентов. Однократное введение вакцины из субъединиц приводило к активизации гуморальных и клеточных факторов иммунитета у мышей и кроликов, повышало их резистентность к экспериментальному заражению и снижало количество клеток ганглия тройничного нерва, несущих геном вируса. — Также рекомендуем «Антигены вируса герпеса. Протеины герпесвирусов.» Оглавление темы «Частная вирусология.»: |
Источник

